

From the point of view of community ecology, soft bottoms are not as well understood as rocky shores. Most experiments using cages tend to alter the sedimentary regime. Further, we do not have a complete understanding of the food base of soft bottoms, particularly for deposit feeders. We cannot say with assurance how much of the non-living organic matter in muddy sediments is used as food. The three- dimensional nature of burrowing also makes it difficult to even see what is going on within the sediment, although many interesting techniques allow us to get around this. We do, however, have much elegant physiological and natural history information about the soft bottom intertidal habitat.

Mud Flat Sampling
Unlike rocky shores, mud flats are far less observable without disturbing the habitat, because many species are infaunal. Typically sediment samples have to be taken and sieved before even major species can be identified. Chemical gradients within the sediment are easily altered by sampling.

Bioturbation
Infaunal animals usually process great amounts of sediment, either by the formation of burrows or by feeding. The yellow arrows point to mounds formed by the burrowing activities of the eastern Pacific intertidal shrimp Callianassa californiensis, whereas the blue arrow points to the rope-like fecal strands of the burrowing polychaete Abarenicola pacifica. The accumulation of sediment on the surface depends strongly upon weather and current strength. During neap tides, these mounds tend to accumulate, reaching heights of several centimeters. During spring tides, when tidal currents are strong and during storm periods, the mounds are eroded away.

Abarenicola pacifica
A head-down deposit- feeding polychaete, A. pacifica maintains a U-shaped burrow, whose bottom may be 20-25 cm below the sediment surface. It feeds upon sediment, and passes rope-like fecal strands to the sediment surface. Jorgen Hylleberg has argued that the feeding activities at the mouth end stimulates bacterial growth, making this species a “gardener.”

Burrowing Shrimp, Callianassa californiensis
Callianassid shrimp are fascinating, though poorly understood ecologically. They form extensive subterranean burrow galleries, which are complex and often interconnected between individuals. The social significance of this is poorly understood, although Darryl Felder and colleagues are now investigating the social and spatial structure of the burrows. Many have investigated the extent of the burrows by pouring marine epoxy resin down the surface holes, and have found complex and lovely arrangments, including burrowing spirals.
Callianassa californiensis is a common resident of muddy sand intertidal flats of the west coast of the United States. It feeds upon paritculate organic matter that it suspends with a feeding current created by the pleopods. The legs collect particles, which are taken by the third maxillipeds and transferred to the mouth.

Callichirus islagrande

Marine biologist removing a cast of shrimp burrow
Photos by (and of) Darryl Felder
An effective way of finding the extent of callianassid shrimp burrows is to pour epoxy resin into the surface burrow holes. Then, one must hand-dig the delicate impregnated burrows (lower left). But the result (lower right) shows the complex burrow structure. This species was collected on a sand flat in east Texas.

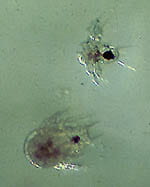
Harapacticoid Copepod Coullana canadensis. Photos by Darcy Lonsdale
Coullana canadensis is a deposit-feeding harpacticoid copepod, common in estuarine sediments of eastern North America. It has a planktonic larva (right), which passes through several planktonic molts before settling to the bottom. The adult is only 1-2 mm in length. Many such smaller macrofauna and meiofauna often swim into the water column and dispersal is often accomplished by adults.

Eel Grass and Sea Lettuce on a Sand Flat, False Bay, San Juan Island, Washington USA
Eelgrass may sometimes grow in spotty abundance as it does in sandy tidal riffles here at False Bay, San Juan Island, Washington. Note the associated thalli of sea lettuce (arrow), which attach to the eelgrass. Word by Levinton and McCartney (1991, Marine Biology) demonstrates that these patches are possible sources of organic matter for sand flat invertebrates.

Eel Grass Bed, Padilla Bay, Washington. Photo by Si Simenstad
Eelgrass, Zostera marina, occurs in dense beds on both the temperate Atlantic and Pacific coasts of the United States, and in European waters. It is a flowering plant, and pollen is transported by water. Plants spread mainly by vegetative growth, through a rhizome system beneath the sediment water interface. It is composed of tough cellulose leaves and there is little or no grazing on the leaves by herbivores, although a number of species can graze the surface film of diatoms and other microorganisms that live on the leaf surfaces.
Eelgrass is very sensitive to light and fails to photosynthesize even at modest reductions of light. As a result, its maximum depth is usually only a couple of meters, even in typically clear water. In recent decades, human additions of nutrients has fueled increased phytoplankton growth, which has made estuarine waters more turbid. Many have argued that this change has been responsible for the precipitous drop in eelgrass in eastern coast United States estuaries. Eelgrass is nearly gone from Long Island Sound, New York and from Chesapeake Bay. Owing to the dependence upon eelgrass during its juvenile stage (see below), the scallop Argopecten irradians has also declined in abundance.

Turtle Grass Bed, Caribbean. Photo by Thomas H. Suchanek
The Atlantic tropics have extensive shallow water beds dominated by the turtle grass Thalassia testudinum. While this grass is also composed of tough cellulose, there are far more grazers capable of feeding successfully. Most particularly the green turtle Chelonia mydas is an effective grazer. It is rather rare these days, however, owing to hunting. The urchin Diadema antillarum is also an effective grazer, but declined greatly in the Caribbean in the early 1980s. Please see the section on coral reefs for information on the formation of bare overgrazed areas in the vicinity of patch reefs.


Bay Scallop Argopecten irradians on Eel Grass Bed, Long Island, New York. Photos by George Rowland
This scallop, Argopecten irradians, lives as an adult on the sediment surface (left) among eastern U.S. eelgrass beds. It is a suspension feeder and greatly prized as food for humans. Adults are hermaphroditic and spawn freely in the water. Larvae are planktotrophic and settle on eelgrass blades. The juvenile scallop remains attached to the eelgrass, several centimeters above the sediment surface (right), which greatly diminishes crab predation.
Adult scallops swim by clapping the valves and forcing water through openings on either side of the hinge. A. irradians swims in a zig-zagging motion by ejecting water alternately through its two jets. Steven Stanley has speculated that the zig-zag motion allows it to negotiate more efficiently through eelgrass blades, but maybe it enables escape from predators.


Mole Crab, Emerita. Photos by Jeff Levinton and Zen Faulkes
Species of Emerita occur on the Atlantic (E. talpoida -left) and Pacific coasts (E. analoga -right) of North and South America. They are swash riders, and move up and down with the tide, in order to maintain position in sediment that is moist, but not turned over by wave action. They have a peculiar oval shape and burrow posterior first with specialized appendages. They suspenson-feed by means of setose antennae, which trap phytoplankton (see right-hand picture, to right).

Mole Crab Burrowing
This is a specimen of the mole crab, Emerita talpoida, burrowing in a wave-washed beach at Atlantic Beach, North Carolina. Work by Olaf Ellers on the swash-riding bivalve Donax variabilis demonstrates that they can sense the pressure of an oncoming wave, which is followed by behavioral responses.
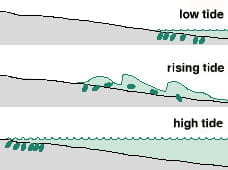
Swash Riding Cycle
Vertical beach migration of swash riders, such as the mole crab Emerita and some species of the wedge clam Donax. Note that animals seek sediments that are neither too dry nor too wave-washed. Animals must be able to sense a pressure difference as the wave approaches and must be able to leave and reburrow rapidly.

Mud snail, Hydrobia truncata
This is a deposit-feeding snail, abundant on temperate and boreal muddy tidal flats of northeastern North America. It occurs in high densities and feeds mainly on surface diatoms. This is a ventral view through a glass plate. Note the broad foot (F) and the proboscis (P), which appresses against the sediment surface during feeding. Fine particles are swallowed, but the snail can scrape diatoms off of the surface of larger sand grains.

The Eastern Mud Snail Tritia obsoleta
This snail is very common in muddy sediments and is a deposit feeder, feeding principally on surface sediment diatoms and particular organic matter. It is often heavily parasitized and is an intermediate host for the parasite causing “swimmer’s itch,” which is a trematode whose dispersal stage burrows into human legs. Often one individual is infested by several species of trematodes. These parasites often invade the gonads, causing reproductive failure.

Bivalve Macoma nasuta, Excavated in Living Position
Species of Macoma are generally deposit feeders, taking up sediment by means of a mobile inhalant siphon, which extends to the surface. Some species, such as Macoma balthica, however, can suspension feed as well. Here you can see a specimen of M. nasuta that was dug up with sediment intact, so you can see the horizontal living position of the shell. This specimen was collected in False Bay, on San Juan Island, Washington.

Intertidal Species of Macoma, Pacific Northwest
In the Pacific northwest there are many species of Macoma. Here are three common intertidal ones that live commonly together on sandy intertidal flats. M. secta tends to occur deeper in the sediment, communicating to the sediment surface by means of long siphons. M. inquinata tends to occur in muddier sediments than the rest.

Amphipod on Sea Lettuce
This amphipod was found living on sea lettuce, Ulva sp. Its color probably derives from the chlorophyll in the sea lettuce. Many seaweeds have chemical defenses. What would be your expectation about the resistance of this amphipod to such defenses? More mobile herbivores?

Burrowing shrimp Upagebia pugettensis
This shrimp lives in deep y-shaped burrows in muddy sand tidal flats of the west coast of North America. It is a suspension feeder.

Burrow entrance, Upagebia pugettensis
The burrow entrances are usually 1-2 cm in diameter and a mound is present, owing to the burrow-maintenance of the shrimp.

Feeding appendages, Upagebia pugettensis
Here is a close-up, showing the feathery feeding appendages, presumably for trapping detrital particles.

Polychaete, Nereis vexillosa
The polychaete Nereis vexillosa burrows in sandy sediments (it also is found among the byssal threads and accumulated detrital material of mussel beds) and feeds on green seaweeds such as Ulva and Enteromorpha.
During the breeding season, the parapodia become better developed and males and females swim into the water to spawn and die. Like other nereids they participate in a “mating dance” and can be seen commonly under night lights in spring off docks on the west coast of North America.

Fiddler Crab. Photo by Richard Zimmer-Faust
There are many species of fiddler crabs, but in all the male has an exceptionally large claw, used for displays to attract females and for intermale combat. Also note the extraordinary stalked eyes, which are excellent for detecting predators and for assessing oncoming females or competing males.

Displaying Male Fiddler Crab, Celuca pugilator, Long Island, New York
Here we see a displaying male Celuca pugilator, right near the entrance to his burrow. Males wave their claws vigorously and successful members of this species copulate with the females in the burrows. After copulation, the female remains in the burrow, incubates the eggs, and then leaves to release larvae into the water.

Fiddler Crab Uca monolifera. Photo by Elliot Norse
Many species of fiddler crabs are brightly colored, which must facilitate recognition. In many places, several species of Uca coexist on the same flat, so accurate species recognition is as important as intraspecific mate recognition.

Horseshoe Crab, Limulus polyphemus
This is a copulating pair of horseshoe crabs, which were photographed on West Meadow Beach, Stony Brook, New York. After copulation, females come onto shore and dig a hole, into which they lay their eggs. Burrowing activity in the intertidal is a major source of mortality for burrowing invertebrates.

Eastern Mud-Flat Oligochaete
This oligochaete, Paranais litoralis is only 1-5 mm long and is a dominant part of the fauna of north Atlantic mudflats. It is hermaphroditic but mainly reproduces by a form of fission. It is a deposit feeder and greatly increases in abundance in late spring and early summer. This population flush is probably caused by deposition of particulate organic matter in spring, but fall deposition can cause a population increase as well, especially when the source is decomposed sea lettuce. When food is exhausted, the populations crash. At the time of the crash a distinct swimming form is believed to allow dispersal to new and better microhabitats that might be richer in food.

Olivella baetica, Sandy West Coasts. Courtesy of Friday Harbor Laboratories
This snail is found in sandy shallow subtidal and low intertidal sediments of the eastern Pacific. It usually lives partially buried and feeds on detrital material. Often, large numbers of shells are washed onto sandy beaches of the west coast of the Pacific northwestern United States.

Cryptic Coloration
Can you spot the fish? On soft-bottom surfaces, there are few crevices within which to hide. Many species of flatfish have chromatophores that permit rapid adjustments of surface color and pattern to the background sediment. Here, a fish has adjusted its color to a mottled sand environment.


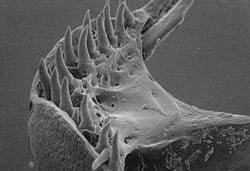
Feeding by a Specialized Sculpin. Photos by Stephen F. Norton
Sculpins are ubiqitous bottom-associated fish that feed on a large variety of marine bottom invertebrates. They can be found commonly in American tidepools and often feed on mobile benthic invertebrates in the same pools. Normally these invertebrates are soft-bodied, but one species in Pacific subtidal bottoms is specialized for feeding on mollusks. Asemichthys taylori can punch holes in the shells of smaller bivalves and snails by means of teeth on a specialized mouthpart (upper left). The puncture holes (upper right) allow digestive enzymes to attack the soft tissues. Bivalves then pass through the gut and are digested (lower x-ray photo).